Promoters for Constitutive and Tissue-Specific Expression of Transgenes in Alfalfa
Deborah A. Samac, USDA-ARS-Plant Science Research Unit, St. Paul, MN
There is great potential for genetically modified alfalfa in the production of industrial raw materials, for new uses such as phytoremediation, and for improving agronomic properties of forage alfalfa. For some traits, the expression pattern desired is constitutive, whole plant expression. For other traits, constitutive expression may be detrimental and expression limited to specific organs such as the nodule, specific cell types such as xylem or phloem, or upon certain environmental cues such as pathogen infection or insect damage may be required. A wide variety of promoters have been described from crop species, but relatively few have been tested for use in alfalfa. Using b-glucuronidase (GUS) reporter gene fusions, my collaborators and I have characterized the expression patterns in transgenic alfalfa of several nominally strong constitutive promoters, several inducible promoters from heterologous species, and a number of alfalfa gene promoters.
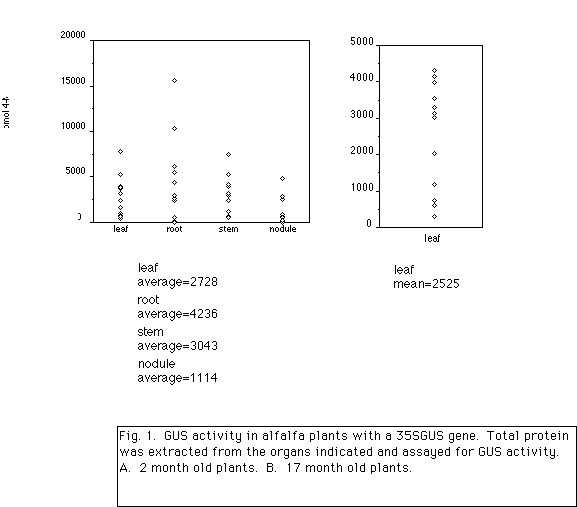
The CaMV 35S Promoter. Undoubtedly the most frequently used promoter in genetically modified plants is the 35S promoter from cauliflower mosaic virus (CaMV). Early studies showed that in virus infected plant cells, this promoter directs high constitutive expression of a viral gene encoding a 35S RNA without requiring additional viral products (Guilley et al., 1982). When the 343 bp of sequence upstream of the protein coding region of the 35S gene is fused to a marker gene such as GUS and stably inserted into a plant chromosome, the gene is strongly expressed in nominally all cells. GUS activity is frequently monitored by histochemical staining in which the substrate X-gluc is incubated with plant pieces (Jefferson, 1987). An insoluble blue dye, formed as a result of enzyme activity, indicates the location of GUS activity. Alternatively, activity may be quantified by extracting protein from plant pieces, incubating with the substrate 4-methylumbelliferyl b-D-glucuronic acid (MUG), and detecting the fluorescent product, 4-methylumbelliferone (4 MU). Frequently, assay results are reported as pmol 4-MU/min/mg protein.
We have utilized the 35S promoter to drive expression of GUS, a number of alfalfa genes in the sense and antisense orientations, as well as genes of bacterial origin. In contrast to the strong activity in other plants, we have observed that the 35S promoter is only moderately active in alfalfa leaves. In leaves of Arabidopsis and tobacco plants containing a 35S-GUS gene, GUS activity is reported to range from 5,000 to 200,000 pmol 4-MU/min/mg protein. As shown in Fig. 1, GUS activity in alfalfa leaves driven by the 35S promoter is 10 to 30-fold lower ranging from 500-7800 pmol 4-MU/min/mg protein. The mean activity in leaves of the plants from Fig. 1 is 2728 pmol 4-MU/min/mg protein. We have also observed rapid declines in GUS activity in alfalfa leaves. While activity in leaves of plants recovered from tissue culture is relatively strong, GUS activity in leaves of older plants, 2-3 months from tissue culture, is often limited to vascular tissue. While some plants retain close to original amounts of GUS activity, others completely lose activity. Further characterization of this gene silencing, which may be a natural plant defense toward foreign DNA sequences, needs to be carried out in order to determine if DNA modification such as methylation of the promoter and/or coding region is occurring, and to establishing the effect of plant genotype on gene silencing. We have observed that not all alfalfa cells express the 35S-GUS gene. In stems, activity is found in the epidermis, chlorenchyma, phloem, and cambium but rarely in xylem or pith cells. In roots and nodules, activity is limited in most plants to only vascular tissues. Clearly, other strong constitutive promoters need to be identified in order to express genes in alfalfa for biotechnological applications.
Expression of Heterologous Promoters in Alfalfa. Numerous tissue specific and inducible promoters have been described from plants. In most cases, the promoter-GUS fusion was tested in the same plant species or within the same plant family. Reports of expression of promoters in heterologous plant species are less common. We have tested several promoters from Arabidopsis and one from potato as promoter-GUS fusions in alfalfa to determine if activity described in the original plant species is retained in alfalfa. In Arabidopsis, the class III chitinase promoter has enhanced expression in roots. Expression is also seen in senescent leaves, hydathodes, guard cells and pollen (Samac and Shah, 1991). Upon infection by foliar fungal pathogens, expression is observed in mesophyll around lesions. A similar pattern of expression is also observed for this promoter in tomato. When the chitinase promoter-GUS gene is expressed in alfalfa, GUS activity is limited to vascular tissue, particularly phloem. Interestingly, upon infection by foliar pathogens, GUS activity occurs around lesions.
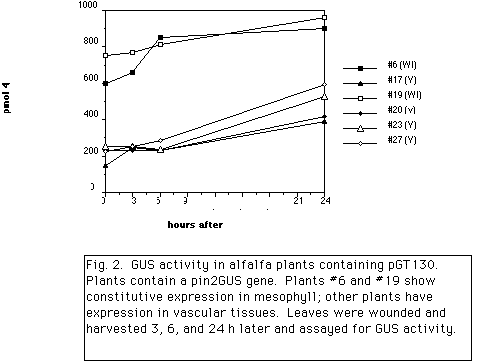
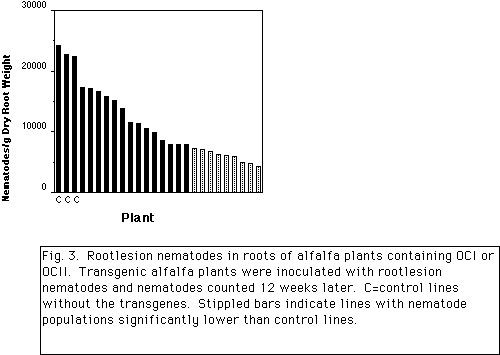
The potato protease inhibitor II (pin2) gene promoter is expressed constitutively in potato root tips. After wounding of leaves by mechanical injury or insect feeding, expression in vascular tissue occurs throughout the plant (Keil et al., 1989). Maximum induction occurs 24 hours after wounding. A similar pattern of expression is observed in tobacco and rice plants containing the pin2-GUS gene (Xu et al., 1993). In alfalfa, two patterns of expression were observed. In one group plants displayed low constitutive activity in leaf and root vascular tissues and root tips, while other plants had low constitutive activity in leaf mesophyll and root tips. As shown in Fig. 2., after wounding leaves, GUS activity in leaves from both groups increased 2 to 3-fold over 24 hours indicating weak induction by wounding. The pin2 promoter was fused to a rice oryzacystatin gene encoding a cysteine proteinase inhibitor and stably integrated in alfalfa plants. Vegetative cuttings of 23 lines were grown in a sand:soil mix in a growth chamber and inoculated with 300 root-lesion nematodes (RLN; Pratylenchus penetrans). After 12 weeks, the number of nematodes in fibrous roots were determined. As shown in Fig. 3., control plants supported an average of 23,120 RLN/g dry fibrous roots. Of the lines tested, 9 lines had significantly fewer nematodes in roots than control plants (P=0.05) with an average of 5,971 RLN/g dry fibrous roots. On average, these 9 lines supported 74% fewer nematodes than control plants, approaching the control obtainable in resistant varieties. The same plants have been evaluated for resistance to the alfalfa weevil but significant control was observed for only one line in 2 of 3 experiments. This difference may be due to the different manner of feeding by the two pests. Experiments to detect OCI and OCII in wounded alfalfa leaves by protein immuno-blotting indicate that the protein in below the limit of detection. Experiments to detect the inhibitors in roots and dried foliar material, and to evaluate resistance to the root-knot nematode are underway.
Expression of Homologous Promoters in Alfalfa. My collaborators and I have characterized the expression of promoters from a number of nodule-enhanced genes central to nitrogen and carbon cycling in alfalfa root nodules including: asparagine synthetase (Shi et al., 1997), aspartate amino transferase (Yoshioka et al., 1999), glutamate synthase (Trepp et al., 1999), and PEP carboxylase (Pathirana et al., 1997). The expression of the promoter-GUS genes were identical or highly similar to mRNA accumulation as determined by RNA blotting and in situ hybridization. For example, the full-length promoter (1277 bp) of the PEP carboxylase nodule-enhanced gene directs expression in nodules as well as in root tips, leaf pulvini, and pollen (Pathirana et al., 1997). In experiments to identify promoter elements involved in nodule-enhanced expression, we found that a 536 bp upstream sequence of the PEP carboxylase promoter is a strong promoter for expression in vascular tissue, particularly xylem. Promoters larger than 536 bp still directed expression in nodules while promoters of 285 and 134 bp directed weak expression in vascular tissues (Pathirana et al., 1997). The xylem-specific expression is relatively novel and may be exploited for expression of specific genes in alfalfa.
We recently isolated and characterized a mitogen-activated protein kinase (MAP kinase) gene from alfalfa with nodule-enhanced expression (Schoenbeck et al., 1999). MAP kinases are a family of protein kinases demonstrated to participate in the cascade of protein phosphorylations by which many diverse signals are transduced within a cell. MAP kinases are expressed in dividing cells but are also associated with response to stress responses such as wounding, drought, cold and fungal elicitors. The alfalfa nodule-enhanced MAP kinase promoter directed GUS expression in the meristematic cells of the root nodule, the root tip, and glandular trichomes. The gene appears to be especially active in developing nodules and roots as X-gluc staining can reveal incipient nodules and root primordia before they have emerged from the root cortex. In stems, expression occurred in the phloem and chlorenchyma. Expression was induced in leaves by mechanical wounding and pathogen infection (Schoenbeck et al., 1999). An identical pattern of expression was observed in Medicago truncatula plants transformed with the same gene construct. When transferred into tobacco, the promoter was active in apical and axillary meristem and in root tips, but activity was lower than in alfalfa (Schoenbeck et al., 1999). These results suggest conservation of transcription factors involved in cell division between alfalfa and tobacco.
Conclusions. There is clearly a need for additional tissue-specific and inducible promoters for expressing genes in alfalfa. However, with the promoters that have been characterized to date, we have ready tools for developing alfalfa plants with unique traits that are capable of growing in new environments, have enhanced nutritional value, and open new markets for alfalfa and alfalfa products.
References
Guilley, H., Dudley, R. K., Jonard, G., Balazs, E., and Richards, K. B. 1982. Transcription of cauliflower mosaic virus DNA; Detection of promoter sequences and characterization of transcripts. Cell 30:763-773.
Jefferson, R. A. 1987. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 5:387-405.
Keil, M., Sanchez-Serrano, J. J., and Wilmitzer, L. 1989. Both wound inducible and tuber-specific expression are mediated by the promoter of a single member of the potato proteinase inhibitor II gene family. EMBO J. 8:1323-1330.
Pathirana, S., Samac, D. A., Roeven, R., Vance, C. P., and Gantt, S. J. 1997. Analyses of phosphoenolpyruvate carboxylase gene structure and expression in alfalfa. Plant J. 12:293-304.
Samac, D. A., and Shah, D. M. 1991. Developmentally regulated and pathogen inducible expression of the Arabidopsis acidic chitinase gene promoter in transgenic plants. Plant Cell 3:1063-1072.
Schoenbeck, M. A., Samac, D. A., Fedorova, M., Gregerson, R. G., Gantt, J. S., and Vance, C. P. 1999. The alfalfa (Medicago sativa) TDY1 gene encodes a mitogen-activated protein kinase homolog. Molec. Plant-Microbe Interact. in press
Shi, L., Twary, S. N., Yoshioka, H., Gregerson, R. G., Miller, S., Samac, D. A., Gantt, J. S., Unkefer, P. T., Vance, C. P. 1997. Nitrogen assimilation in alfalfa: Isolation and characterization of an asparagine synthetase gene showing enhanced expression in root nodules and dark adapted leaves. Plant Cell 9:1339 1356.
Trepp, G. B., van de Mortel, M., Yoshioka, H., Miller, S. S., Samac, D. A., Gantt, J. S., and Vance, C. P. 1999. NADH-Glutamate synthase (GOGAT) in alfalfa root nodules: Genetic regulation and cellular expression. Plant Physiol. 119:817-828.
Xu, D., McElroy, D., Thornburg, R. W., and Wu, R. 1993. Systemic induction of a potato pin2 promoter by wounding, methyl jasmonate, and abscisic acid in transgenic rice plants. Plant Molec. Biol. 22:573-588.
Yoshioka, H., Gregerson, R. G., Samac, D. A., Hoevens, K. C., Trepp, G., Gantt, J. S., and Vance, C. P. 1999. Aspartate aminotransferase in alfalfa nodules: Localization of mRNA during effective and ineffective nodule development and promoter analysis. Mol. Plant-Microbe Interact. 12:263 274.
{kind=link}
{kind=link}
{kind=link}